1Assistant Professor, Department of Zoology, Government Degree College, India.
2Research Scholar, Government Degree College, India.
Arti Sharma
Email: arti.biochem05@gmail.com
Received : Dec 16, 2024 Accepted : Jan 20, 2025 Published : Jan 27, 2025 Archived : www.meddiscoveries.org
Microalgae is one of the best sources of renewable energy production, such as biofuels. The production of biodiesel from microalgae has several advantages, including the high productivity of lipid and the possibility of cultivating them on marginal land. One of the challenges in using microalgae for biodiesel production is the complexities process of lipids extraction by organic solvents followed by transesterification. The aim of this work is to optimize this process by a single extraction and conversion step. The reaction was carried out for different parameters such as; various oil to methanol ratios, concentration of catalyst, temperature and time reaction. The algal biodiesel samples were analyzed with Gas Chromatography Mass Spectrometry (GC-MS) and fourier transform infrared spectroscopy in this study can be considered as potential feedstock for biodiesel production to fight the future energy crisis. Biodiesel production from microalgae is being widely developed at different scales as a potential source of renewable energy with both economic and environmental benefits. Although many microalgae species have been identified and isolated for lipid production, there is currently no consensus as to which species provide the highest productivity. Different species are expected to function best at different aquatic, geographical and climatic conditions. In addition, other value-added products are now being considered for commercial production which necessitates the selection of the most capable algae strains suitable for multiple-product algae biorefineries. Microalgae isolation and selection for traits that maybe most relevant for commercial biodiesel production.
Keywords: Biodiesel; Fatty acid methyl ester; Marine microalgae; Transesterification.
The primary producers of oxygen in aquatic environments are algae, especially planktonic microalgae. These microorganisms are widely distributed in nature and have adapted to different environments with great diversity in size, morphology, life cycle, pigments, and metabolism. About one half of global photosynthesis and oxygen production is accomplished by marine microalgae. They play an important role in CO2 recycling through photosynthesis, which is similar to higher plants in O2-evolved systems (PSI and PSII). Research in microalgae has been carried out not only on physiological aspects but also to develop production of useful biomaterials. The advantages of their utilization in production are their ability to convert CO2 to useful materials through photosynthesis and their ability to grow in natural environments under inorganic conditions. For example, marine microalgae can be cultivated using seawater, CO2, and sunlight. Recent developments in the biotechnology of microalgae have been focused on their production of useful materials applicable to the cosmetic and medical fields. Genetic modification and molecular tools have been developed mainly in eubacterial microalgae, cyanobacteria (blue-green algae). In contrast, genetic modification has been only gradually applied to eukaryotic microalgae. Recently, whole genome sequences and EST analyses have been performed in marine strains. The elucidation of genome wide information may help in the development of new biotechnological applications using microalgae. In this chapter, we review the useful applications of microalgae for genetic engineering, cultivation technologies, and CO2 fixation as follows:
1. Production of useful chemicals by marine microalgae
2. Metabolic engineering of marine microalgae
3. Microalgal mass cultivation technologies
4. CO2 fixation using microalgal cultures in industry
Microalgae are photoautotrophic organisms with different phyla such as Cyanophyta, Chlorophyta, Rhodophyta, Haptophyta, Streptophyta, and Heterokondophyta. Primary and secondary metabolites produced in microalgae are complex organic compounds accumulated by microalgae with the help of H2O, CO2, and energy from sunlight. They are adapted to different environmental stress conditions that extend their growth ranging from freshwater to extreme salinity. They can survive on moist, black earth, and even on desert sands, extending up to clouds. Their large spectrum of existence leads to varieties of chemical compounds with a myriad of properties and represents a remarkable opening to discover novel metabolites, or to produce available metabolites at lower costs. Microalgae possess metabolic versatility and plasticity that are commended in physiological states. Their secondary metabolism can be effortlessly activated by externally applied stresses. Microalgae are deemed to be cosmeceutical with their effortless response to stress. Cosmeceuticals are products with biologically active ingredients with medical or drug like advantages, which aim to improve the structure, morphology, and appearance of skin. Polysaccharides are the active compounds in microalgae. They have potential application in the prevention of blemishes, damaged skin repairing, and inhibiting inflammation process (e.g., genus chlorella). Hence, they are used to produce thickener moisturizers and gelling agents. Numerous other bioactive substances in microalgae accelerate healing process and maintain skin moisture. The quest for these biologically active molecules to treat metabolic disorders and immune ailments drives the way to harvest bioactive compounds from microalgae. The easiness of micro-algal cultivation benefited from using its metabolites in biological applications related to health.
Microalgae are considered as sustainable source of biologically an equal contribution active compounds like biofuels, foods, feed, and pharmaceuticals (Figure 1). Micro algae have 20%-40% higher productivity when compared to oil crops. Some of them can build up to 80% dry lipid biomass weight. Another application of biologically active molecules in microalgae is as nutraceuticals, which focus on saying “Prevention is better than cure.” Microalgal nutraceuticals include compounds like phycocyanin, astaxanthin, Beta carotene, fucoxanthin, lutein, lycopene, phycobiliproteins etc. Microalgae are creators of biomolecules that are relevant in diverse domains. Here in this chapter, we will focus on bioactive compounds and pharmaceuticals from microalgae.
Microalgae sampling
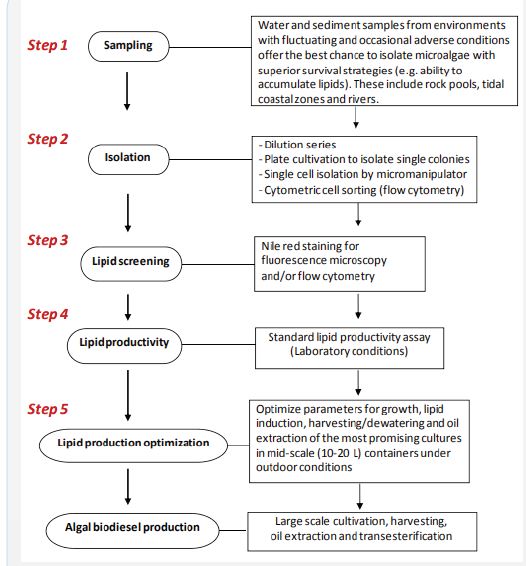
Microalgae occur in most natural environments, including water, rocks and soil; interestingly, they also grow on and inside other organisms. Their main habitats are freshwater, brackish and marine ecosystems. Microalgae can be found and collected not only in common aquatic ecosystems such as lakes, rivers and oceans, but also in extreme environments such as volcanic waters and brackish waters. Local microalgal species should be collected because it can be expected that they have a competitive advantage under local geographic, climatic and ecological conditions. The hypothesis suggests that water and sediment samples from aquatic environments that undergo fluctuating, sometimes unfavourable, conditions provide a greater likelihood of isolating microalgae that accumulate high lipids [8].
Microalgae isolation
Isolation is a necessary process to obtain pure cultures and presents the first step towards the selection of microalgae strains with potential for biodiesel production. Traditional isolation techniques include the use of a micropipette for isolation under a microscope and cell dilution followed by cultivation in liquid media or agar plates. Single cell isolation, based on traditional methods from the original sample is time-consuming and requires sterilized cultivation media and equipment. but the result of this elaborate process is always a pure culture that is usually easily identifiable. Another approach in the laboratory includes the enrichment of some microalgae strains by adding nutrients for algal growth. The most important nutrient sources for algal growth are nitrogen and phosphate.
Strain identification
In the laboratory includes the enrichment of some microalgae strains by adding nutrients for algal growth. The most important nutrient sources for algal growth are nitrogen and phosphate. Some particular algae species may require trace minerals for their growth (e.g., silicon for diatoms). Soil water extract is an excellent source of nutrients for algae growth at this stage because this medium is easy to produce and satisfies nutrient intake of many algae strains. An automated single cell isolation method that has been developed and widely used for cell sorting is flow cytometry. This technique has been successfully used for microalgae cell sorting from water with many different algae strains, primarily based on properties of Chlorophyll Autofluorescence (CAF) and Green Autofluorescence (GAF) to distinguish algae species such as diatoms, dinoflagellates or prokaryotic phytoplankton. Unlike for many agricultural crops, a targeted selection and domestication of microalgae strains is still in its infancy, while technology to economically grow microalgae with high lipid content is still being developed [7].
Microalgae cultivation, biomass and total lipids measurements
Microalgae strains with potentially high lipid content (e.g., as determined by Nile red staining) need to be cultured to increase biomass and directly compared to each other in larger cultivation systems to assess their potential as biodiesel feedstock. Initial tests of the most promising algae strains usually are carried out at laboratory-scale using culturing flasks and other vessels, such as hanging bags, under well-defined growth conditions. The test should follow a standard protocol over a certain growth period to allow direct comparisons between strains in terms of growth rate and lipid accumulation (“lipid productivity”). It should be noted though that a standard assay does not take into consideration the potential of certain microalgae under carefully optimized conditions. An example of this assay is the following that is routinely used by our laboratory to compare lipid productivity: Pure (but not axenic) algae strains are cultured in F/2 medium (fresh or seawater) until near stationary growth occurs (less than 20% growth in 24 h as determined by cell counts). An inoculum of 5 mL of this culture is then used to start growth in 20 mL fresh F/2 medium exactly at 2 h after the start of the light cycle. The culture is then grown and monitored by cell counting for 7 days after which medium is replaced with nutrient-free water [3]. Nutrient starvation is conducted after that for another 2 days of cultivation to test the potential for rapid TAG accumulation. In addition, biomass is collected at the end of the experiment for lipid content analysis. This assay is useful for screening of growth and lipid producing capacity of microalgae, leading to selection of potentially useful strains.
The best candidate strains with potential for biodiesel production should then be used to optimize parameters for rapid growth, lipid induction, harvesting/dewatering and oil extraction. While most of these parameters are typically optimized under small-scale laboratory conditions, it seems advisable to move towards larger size outdoor cultivation conditions as soon as possible, as these are typically quite different [2]. Parameters, such as salinity, nutrient composition, pH and cell density can be controlled to some extent, but other factors such as temperature, irradiation and the co-cultivation of other organisms are much harder to control under outdoor conditions. it is advantageous to isolate and screen a large number of local microalgae strains and test these under mid-scale outdoor conditions as soon as possible to be close to conditions that maybe expected for large-scale cultivation. Figure 2 provides a step-by-step overview of how microalgae may be rapidly isolated and selected for larger scale biodiesel production. At larger scale, two popular cultivation systems have been used for microalgal biomass and lipid production: open raceway ponds and closed photobioreactors. At present, open pond systems, especially large raceway ponds, are much more widely used, but bear the risk of attracting competing algae, grazers or viruses. Although minimizing the cost of algae farming is one of highest priorities to achieve commercial algal biodiesel production, both systems require optimization of complex factors that satisfy high level production cultivation.
Lipid extraction and Trans-esterification reaction
Lipid extraction is usually carried out using three different techniques (a) Bligh and Dyer method (b) Folch method (c) Soxhlet extraction method.
Bligh and dyer method: In this method, two-phase systems are created, and the lipid is fractioned out from the chloroform phase of the solution. Bligh and Dyer’s method modified with a gravimetric approach has been utilized for the lipid extraction from C. regularis and optimum lipid quantity was obtained. Lipid extraction Bligh and Dyer method, in the presence of methanol: chloroform (1:2, v/v) as a solvent mixture. Further, transesterification was carried out using methanol as the sole solvent [4].
Folch method: Folch method, similar to Bligh and Dyer method, is a static method used for lipid extraction. It utilizes chloroform and methanol as extraction solvents. Lipid from Monoraphidium sp. T4X cultivated in BG11 medium was extracted using the Folch method and the lipid content was found to be 18.42±0.4%. Lipid extraction from dried biomass of Folch method, using methanol: chloroform (2:1, v/v) as a solvent mixture. Further, transesterification was carried out using methanol as the only solvent.
Soxhlet extraction method: This is a dynamic method of lipid extraction where conventional solvents such as methanol, hexane, and ethanol are used and the process is accompanied by the use of Soxhlet apparatus. Soxhlet extraction process has been used for lipid extraction from M. decolor dry biomass using ethanol and hexane giving 23.78±0.68% and 12.71±0.2% lipid yield respectively. whereas the Folch and Bligh-Dyer method requires 50 mg to 1 g of the same. Soxhlet apparatus helps in more efficient lipid extraction as it is a dynamic process whereas the other two being static extraction methods may result in low lipid extraction [5].
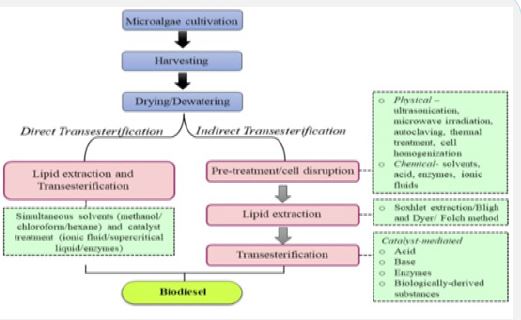
Transesterification is the process of converting one molecule of triglyceride to three molecules of ester and glycerol, primarily carried out using an acid or a base catalyst. Generally, methanol is used as the alcohol for the transesterification process as it is cheap and produces low molecular weight alkyl esters called Fatty Acid Alkyl Ester (FAAE) or biodiesel. The amount of FAAE obtained, or the rate of conversion is affected by the methods used for lipid extraction and determination. Lipids containing low levels of transesterifiable lipids also result in low FAAE yield. There are two broad ways of carrying out transesterification, viz., direct and indirect transesterification process. Direct transesterification is carried out without disrupting microalgal cells with wet microalgal biomass whereas indirect transesterification involves cell disruption and lipid extraction as the preliminary steps before performing the transesterification process.
Direct transesterification
Direct transesterification or in situ transesterification is a cost-effective approach as it utilizes the wet microalgal biomass, thereby reducing the pre-treatment and lipid extraction costs. It is a combination of two stages, viz., lipid extraction and biodiesel conversion using catalysts that intensify reactive separation. It can be carried both with and without the catalyst. Various microalgae have been subjected to direct transesterification for optimum FAAE/ biodiesel yield. Direct supercritical methanol transesterification facilitates lipid extraction and conversion to biodiesel in a single step. The process requires high temperature (240-290°C) and pressure (9.3-30 MPa). The wet microalgal biomass contains a high amount of water and free fatty acids. This aids the supercritical conversion of lipids to biodiesel. The end product does not undergo the purification process if catalysts are not added.
Indirect transesterification
Indirect or conventional transesterification processes convert lipids into alkyl esters using catalysts at 60 to 70°C. First, cell disruption is carried out followed by lipid extraction using various solvents like methanol, chloroform, and n-hexane. The extracted lipids are then transesterified using a homogenous or heterogenous acid, base, or biocatalyst. Catalyst selection is crucial for enhanced biodiesel production as it affects the transesterification process. The main function of the catalyst is to reduce the amount of activation energy required for the reaction to be carried out. There are three broad categories of catalysts: acids, bases, and biocatalysts. Acids and bases are the conventional catalysts whose activity depends upon the fatty acid content of the algal oil. In an acid-mediated reaction, protonated carbon of the carbonyl group of triglycerides reacts with the –OH from the solvent (methanol/ethanol), creating a tetrahedral intermediate product. Whereas in a base-mediated reaction, the nucleophilic alkoxide is created by the –OH group from methanol/ethanol that reacts with the electrophilic part of the -C=O group of triglycerides [6]. The conventional catalyst (acid/base) can be further classified as a homogenous and heterogenous catalyst. Further, the catalyst activity may be regulated by modifying the reaction time, working conditions (temperature and pH), and activation assisted with saturated solutions such as Ca(OH)2.
GCMS
Gas Chromatography-Mass Spectrometry (GC/MS) is one of the most frequently used instruments in metabolomic analysis because of the relatively low cost, reproducibility, stability, and convenient data processing compared to other instruments such as Liquid Chromatography-Mass Spectrometry (LC/MS) or Nuclear Magnetic Resonance (NMR). Previously, GC/MS was commonly used to analyze volatile compounds because of its ability to examine gas-phase samples. However, the development of various derivatization methods has enabled researchers Gas Chromatography-Mass Spectrometry (GC/MS) is one of the most frequently used instruments in metabolomic analysis because of the relatively low cost, reproducibility, stability, and convenient data processing compared to other instruments such as Liquid Chromatography-Mass Spectrometry (LC/MS) or Nuclear Magnetic Resonance (NMR). Previously, GC/MS was commonly used to analyze volatile compounds because of its ability to examine gas-phase samples. However, the development of various derivatization methods has enabled researchers to analyze low molecular weight non-volatile compounds. This advancement brings a new perspective to the potential application of GC/MS in the metabolomics world. High reproducibility of retention time and mass spectra are the main advantages of GC/MS. These advantages have been made possible due to the development of GC/MS over the past decades into a mature technology. High reproducibility of retention time is achieved by calculating the retention time index of the peaks detected in n-alkene or fatty acid methyl ester solution and comparing it to the databases that is already available online, including the Fiehn library, Golm Metabolome Database, MassBank, Wiley Database, and National Institute of Standard and Technology (NIST). In addition, mass spectra identification is possible because of the standardized method of electron ionization to 70 eV; thus, the online library can be utilized freely as long as the analytical settings are the same. With the combination of derivatization process that enabled the analysis of non-volatile compounds in addition to the volatile compound and GC/MS advantage that are robust and highly reproducible, it is not surprising that GC/MS is quickly emerging as a researcher’s preferred choice of instrument. However, in the application of GC/MS for non-volatile compound analysis, the need for derivatization can become a drawback of this technology. Derivatization may lead to unstable derivatization of amino acids, resulting in two peaks and the production of several sugars peak due to their geometric isomers, thus preventing accurate quantification and identification, or producing additional reactions such as pyroglutamate from glutamate. This drawback implies that researchers who work with GC/MS need to take these changes into consideration to correctly understand the mechanism in their samples.

Fourier transform infrared spectroscopy
Fourier-Transform Infrared (FT-IR) spectroscopy is a vibrational, non-destructive method which allows the simple and reproducible analysis of the tissue with only small amounts of material. It is based on the change in the dipole moment in a molecule, working on the principle of vibrating molecular bonds and the resulting absorption wavelengths, which depend on the involved atoms and strength of intermolecular interactions. Despite providing specular results as RS, FT-IR mainly deals with non-aqueous samples, because of the strong absorption bands of water. Moreover, compared to RS, FT-IR has higher sensitivity but it does not provide the possibility of confocal resolution, and its lateral resolution is 10-20 μm (compared to 1-2 μm of Raman spectroscopy). Nonetheless, it has been widely used for diagnostic purposes thanks to the possibility to obtain quantitative information from the biological sample and to the fast acquisition (few minutes). These properties make this technique a suitable tool for the Point-of-Care (POC) analysis. Under the economic point of view, commercially available instrumentation is usually less complex and expensive than the corresponding Raman spectrometers. In the EV field, FT-IR was used as fast and simple characterization method of EV isolated from cultured cells. The IR spectra were used to gain general information concerning the molecular constituents and the structures of EVs and apoptotic bodies after drying the sample. Results demonstrated that the typical IR spectra of apoptotic bodies resemble those of the parent cells, whereas EVs differ from the cell source due to changes in the spectral features of protein secondary structure. Furthermore, Mihály and colleagues introduced a new parameter, i.e. the spectroscopic protein to lipid ratio, to classify different pre-isolated EV subpopulations. IR spectroscopy was demonstrated to represent a reliable, fast and relatively cheap screening approach for EV isolation which requires only minimal amount of sample and no complex sample preparation procedures, and it was proved to be able to assess differences in the purity level of EV preparations and differences in EV subtypes (large vs small EVs). Although firstly proposed for research purposes to efficiently classification of EV subpopulations, in 2016 another research group deposited a patent that suggested FT-IR for diagnosing, prognosing and monitoring pathophysiological states, like cancer. Results related to the analysis of EVs isolated by a combined protocol of ultrafiltration and size exclusion chromatography from the supernatant of cultured breast cancer cells were reported. Spectral data demonstrated that three different cell lines owed distinct biochemical features related to their cellular origin and possibly by the extension to cancer cells and to the tumor stage. FT-IR was also demonstrated to have potentialities in the identification of the biochemical fingerprint of blood and salivary EVs. Compared to blood EV results that were characterized by significant contribution of protein components, maybe due to the selected isolation procedure, IR spectra of salivary EVs revealed differences in multiple biomolecules and in particular differences were observed in the peak at 1072 cm−1, corresponding to absorbance bands of phosphate bonds within nucleic acids. Such observation made authors conclude that the content of nucleic acids in cancer EV can be lower than that from healthy EV, although further investigation is needed [1]. Collectively, FT-IR can be considered as an alternative and/or a complement to other methods used for fingerprinting EV subpopulations, including other spectroscopic methods, such as RS, as it was demonstrated to be able to identify general features shared between several EV subtypes or specific to one sub-type of EVs. Still, the identification of the proper isolation method is crucial for the downstream application of IR data.

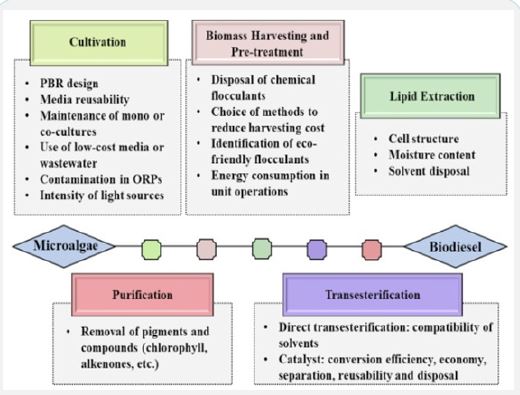
Biodiesel production from microalgae has been divided into different stages viz., cultivation and maintenance of microalgae, biomass harvesting, cell disruption, lipid extraction and finally lipid to biodiesel conversion (transesterification). Each step offers certain challenges such as high-power consumption, use of toxic chemicals and cost-ineffective processes (Figure 6). Extensive research is required to establish a standard method for efficient biodiesel production. Challenges involved at each stage of production process discussed further in this section.
In this study, 14 isolates of marine microalgae were obtained from the different samples collected in the ocean. The purification of the isolates was carried out using repeated streaking on the solid BG11 medium. The purification allowed monoalgal culture harvested on BG11 liquid medium. The BG11 medium was very useful in this work for the isolation of marine microalgae belonging to different groups. Microalgae isolation from seawater samples require the use of several types of media containing concentrations of many essential nutrients in order to allow the growth of the majority of microalgal species present in the samples. Common examples of media used for marine algae are Black Sea medium, ES Medium, ASP Medium, Aquil medium, Allen’s Cyanidium, CCAP Artificial seawater, Chry medium, ESAW Medium, ESM Medium, f/2 medium, K Medium, L1 Medium, MNK Medium, Pro99 Medium, SN medium etc.
In this study marine microalgae are tiny single-celled algae found in ocean waters. They play a crucial role in various biological process and are vital to the earth’s ecosystem. Biofuel is a type of renewable energy source derived from biological material such as plants agriculture waste and other organic matter. It serves as an alternative to fossil fuels and can help reduce greenhouse gas emissions.