Faculty of Health and Life Sciences, Department of Pharmaceutical Biotechnology, Portland Laboratory in De Montfort University in Leicester, United Kingdom.
Al Baraa Akram
Email: albraaakram94@gmail.com
Received : Feb 18, 2026 Accepted : Mar 19, 2026 Published : Mar 26, 2026 Archived : www.meddiscoveries.org
In nematodes, caspase activation can cause apoptotic cell death during developmental cell death, which is then followed by adjacent cells phagocytosing the dead cell. Phosphatidylserine exposure on the cell surface is linked to apoptotic cell death and can serve as an eat-me signal for phagocytosis. However, for certain nematode cells, caspase activation is sufficient to cause phosphatidylserine exposure for surrounding cells to phagocytose the living cell, which results in phagocytosis-induced cell death.
Although there is evidence that opsonin’s and lectin receptors that recognize mannose residues on the prey surface, as is the case with mammalian phagocytes, are used by phagocytic protozoa to recognize and consume their prey, relatively little is known about this process. On the other hand, the G-protein-coupled receptor FAR1, which binds sugars on bacterial LPS, is used by the amoeba Dictyostelium disodium to facilitate both chemotaxis towards and phagocytosis of bacteria.
Keywords: Phagophobia; Eat-Me Signals; Find-Me Signals.
Caspases can cause apoptotic cell death during developmental cell death in worms, which is followed by nearby cells phagocytosing the cell corpse [1]. Phosphatidylserine exposure on the cell surface is linked to apoptotic cell death and can serve as an eat-me signal for phagocytosis. Nevertheless, in certain nematode cells, caspase activation is sufficient to cause phosphatidylserine exposure and phagocytosis of the living cell by nearby cells, which leads to cell death via phagocytosis [2,1].
Numerous cell types can reverse caspase activation and phosphatidylserine exposure; for instance, inflammatory phagocytes can cause sub-toxic caspase activation and reversible phosphatidylserine exposure on target cells, which results in phagocytosis-induced cell death [3]. Therefore, apoptotic cell death is not always indicated by apoptosis indicators such caspase activity. Additionally, in certain situations, phagocytosis induced cell death may result in apoptosis of the engulfed cell, whereas in other situations, sub-lethal stimulation of apoptosis pathways may ultimately result in cell death. But there is still a clear difference between apoptotic cell death and phagocytosis induced cell death: blocking phagocytosis would stop the former but not the latter.
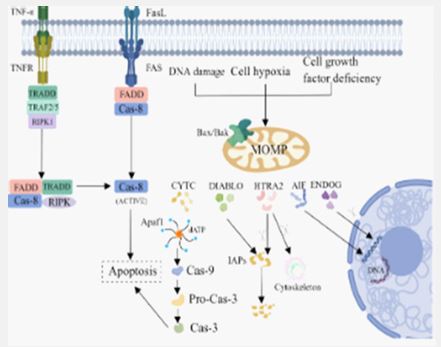
Although there is evidence that opsonin’s and lectin receptors that recognize mannose residues on the prey surface, like those found in mammalian phagocytes, are used by phagocytic protozoa to recognize and consume their prey, relatively little is known about this process [4]. But to chemotaxis and phagocytose bacteria, the amoeba Dictyostelium disodium needs the G-protein-coupled receptor FAR1, which binds sugars on bacterial LPS [5].
It can be challenging to image phagophobia because it necessitates serial or continuous imaging of the same cells over time; phagophobia is a relatively uncommon occurrence that happens rapidly; and it can be challenging to determine whether the cell being phagocytosed is alive or capable of living if phagocytosis was avoided. Nevertheless, various methods have been advised. When phagocytosis was inhibited, the neurons remained alive, and video imaging of active microglia and neurons in culture revealed microglia devouring intact neurons with uncondensed nuclei [6]. Without any signs of apoptosis or DNA damage, retinal microglia were seen to engulf the stomata of intact photoreceptors (rod cells) in vivo. When phagocytosis was inhibited, the photoreceptors continued to function [7].
By staining the cells with JC-1, which turns the mitochondria red when polarized and green when depolarized, the viability of lymphoblasts during phagocytosis was observed. The addition of anti-CD47 antibodies caused macrophages to engulf polarized (viable) lymphoblasts, which subsequently depolarized, signifying that viable cell had been phagocytosed [8]. Similarly, using the dye DilC1(5) to track mitochondrial polarization in the oligodendrocyte precursor cells, which were caspase 3-negative when ingested by microglia, the engulfment of viable oligodendrocyte precursor cells by microglia was verified in brain slices [9]. Cyst cells phagocytosed sperm progenitors before any DNA damage occurred, and when phagocytosis was stopped, the progenitors remained alive, according to imaging studies conducted on Drosophila testis utilizing Lysol Tracker to track lysosomal acidity and DAPI and TUNEL to track DNA condensation and damage [10,11]. Therefore, it is conceivable to image phagophobia, but doing so in vivo is still difficult.
How is phagocytosis induced or inhibited by target cells?
By releasing find-me signals, exposing eat-me signals, binding opsonin’s, and eliminating don’t-eat-me signals, cells can cause phagocytes to phagocytose them. Prior to phagocytosis, phagocytes are chemoattracted to the target cell by find-me signals, which are chemotactic chemicals generated by the target cell. The complement components C3a and C5a, formyl peptides, CX3 C-Chemokine Ligand 8 (CXCL8; also called IL-8), sphingosine-1-phosphate, and the nucleotides ATP, ADP, UTP, and UDP are among them. These mediators work together to activate different chemotactic receptors on phagocytes [12].