1Odessa National Medical University, Odessa, 65082, Ukraine.
2Healthcare Analysis & Forecasting, Wantage, UK.
Andrey Ponomarenko
Email: aponom@hotmail.com
Received : May 01, 2026 Accepted : May 22, 2026 Published : May 29, 2026 Archived : www.meddiscoveries.org
Immunity is a basic feature of life. Some immunological mechanisms are not completely understood. Burnet’s Clonal selection Theory barely explains how a limited number of immune cell clones can ensure immunity to numerically superior antigenic determinants, and how strict timing of immune reactions can be achieved. To solve existing contradictions, we propose a virus-dependent theory of acquired immunity where all the diversity of antigen receptors on the surface of immune cells is not pre-existent but arises from individual immune system encounters with a restricted number of antigens. Adaptive immunity results from multiple atypical latent virus infections with reactivated specific subpopulations of Endogenous Retroviruses with these stages: 1) antigen-dependent reactivation of specific clones of the defective HERVs; 2) targeted infection by reactivated HERV clones or transfection of genes to naïve immune cells causing transformation into Ag-specific T- and B-cell clones accompanied by secretion of specific HERV proteins from primed B-cells as antibodies or their manifestation on cell membrane of primed T-cells. This mechanism resembles Burnet’s Clonal Selection Theory but implements antigen recognition on the molecular level by endogenic virus-like particles (HERV and other retrotransposons), instead of cell clones. This explains strict timing of immune reactions and ability to respond to any new antigen. The Virus-dependent mechanism is metabolically beneficial because it does not imply waste of energy and resources (proteins, lipids, etc.) on the creation of billions of immune cells specifically targeted against all possible antigens, most of which will never be encountered during a lifespan.
Keywords: Acquired immunity; Endogenous retrovirus; Defective interfering viral particles; Clonal selection; CRISPR.
Those who study biology usually admire Nature’s ingenuity and thrift. For example, Nature only uses 4 “characters” in a DNA molecule: adenine (A), guanine (G), thymine (T) and cytosine (C) to encode structure of all known proteins in any living being on the Earth. Nature preserves all efficient mechanisms and principles found at the very early stages of bio-evolution and uses them in all further evolution stages. For instance, the central pathways of metabolism, such as glycolysis and the citric acid (Krebs) cycle, are present in all three domains of living things: bacteria, archaea and eukaryotes, and were present in the last universal common ancestor [1,2]. Same or similar proteins are often present in most living beings from bacteria to human cells and implement similar functions. For example, cryptochromes function as photoreceptors and/or participate in DNA reparation or DNA regulation (circadian clocks) in bacteria, plant and human cells [3]. Meanwhile, when we consider currently accepted mechanism of acquired immunity: the Clonal Selection Theory, proposed by Macfarlane Burnet [4], the feeling of incoherence and wastefulness of Nature is present. We believe that our hypothesis of virus-dependent mechanisms of adoptive immunity [5,6] better matches the principles of prudence and evolutionary conserved similarity of main biological functions. Besides, proposed hypothesis allows avoiding some discrepancies and questions arising during examination of the Burnet’s Theory. It is necessary to mention that there are some other scientists who adhere to the idea of the virus-dependent origin of adaptive immunity [7,8]. Luis Villarreal greatly contributed to this field; he is considering whole human evolution from the perspective of the great HERV colonization [9].
Questions arising from evaluation of the Clonal Selection Theory
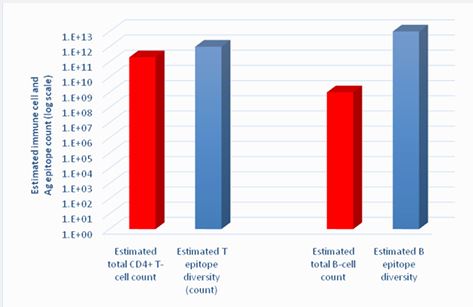
Statistics and probability theory - related questions: According to Macfarlane. Burnet’s Theory, the immune response can be initiated when an antigen in its original or pre-processed form directly encounters corresponding pre-existing receptor on pre-existing B- or T-cell. Estimated number of potential antigenic epitopes in biosphere is enormous and exceeds 1x1012 [10-12]. Thus, the Clonal Selection -based immune response requires pre-existence in each human or animal body at least 1x1012 variants of potentially clonogenic B- and T-cells, each of distinct antigen specificity. Estimated total number of T- and B cells in a healthy human is significantly smaller, not to mention even smaller quantity of specific clones these cells can form (Figure 1) [13,14]. The only explanation of the immune system functionality in these conditions is cross-reactivity of T-cell receptors and affinity maturation in B-cell receptors [13,15]. If not discuss the hazard of autoimmune reactions in conditions of significant T-receptor cross-reactivity, just imagine the probability and time necessary for accidental direct encounters of an antigenic epitope with a complementary T-cell receptor. It looks less feasible than the probability of catching a criminal among all the population on the Earth when several policemen only have his fingerprints and no photo. It is even more frustrating that billions of other T-cells - “policemen” could thousands of times “check fingerprints” of that criminal, but do not recognize him, because of incompatibility of their antigen receptors. Not to mention such wastefulness when billions of immune cells will never ever fulfil their function because an animal never encounters antigens of corresponding specificity.
Also, there is low probability of achieving such a huge amount of different antigenic specificities as a result of accidental genomic rearrangements, because of regular patterns of distribution of a random variable known from statistics: see (Figure 2). This means that to achieve 1x1010 different specificities of pre-existing antigenic receptors, the total number of immune cells in the body must be significantly bigger than it is in reality. Besides, because of significant difference in quantity of immune cells bearing each specific antigen receptor, the time of immune response to various antigens would significantly differ.
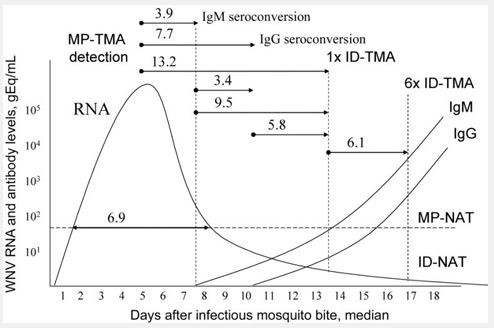
Inconsistency with the strict timing of immune reactions: Now, when we realize the complexity and various statistical probabilities of the direct encounter of an invading antigen with complementary antigenic receptors, could we imagine that this process: the antigen recognition and initiation of the specific immune response - has so strict timing (Figure 3)?
Inconsistency with formal logic: A mouse is 3000 times smaller than human (Figure 4); it contains thousands of times fewer immune cells. Does it mean that the immune system in mice can respond to thousand times less quantity (diversity) of antigens in comparison with human’s immunity? What about blood donors? Does their immune system constantly exhaust its ability to recognize definite amounts of antigens after every blood donation?
Scientific findings that don’t fit the clonal selection theory: Some authors suggest that the immune system practices molecular selection of receptors in addition to clonal selection of lymphocytes [18]. This idea is a serious challenge to Burnet’s theory, because he stated that “The clonal selection hypothesis could be disproved by showing that cells of a pure clone could, by appropriate manipulations, be induced to produce any one of a variety of antibodies” [4]. Research suggest that random mutation and selection as envisioned by Burnet cannot entirely account for B cell receptor editing: B cells can be “manipulated” by an antigen to produce “any one of a variety of antibodies”:
- First, in developing B cells, antigen receptor binding to self antigen induces new V(D)J recombination [19].
- Second, binding to self-antigen induces replacement of the autoreactive receptors (genes) by non-self-reactive receptors [20].
Therefore taking into account all existing contradictions, is it possible that another, less resource-wasteful and more reliable mechanism of acquired immunity exists?
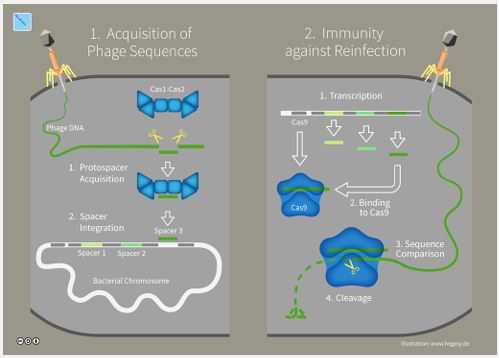
Mechanism of acquired immunity found in bacteria: We proceed from the fact that Nature often uses previously elaborated approaches in its further and more complicated creatures. The majority of fundamental life functions are universal and possess similar features in most living beings (i.e., structure of DNA, RNA, transcription, translation, DNA reparation, metabolic processes, etc.). Therefore, feasible way to decipher immunological mechanisms in animals could be evaluation of similar protective functions in unicellular life forms, which appeared earlier on the Earth, for example - in bacteria. Fortunately, a mechanism of anti-virus (anti-bacteriophage) defense in bacteria and archaea based on CRISPR-Cas complex was recently discovered [21,22]. CRISPR (clustered regularly interspaced short palindromic repeats) are specific DNA sequences found in the genomes of prokaryotic organisms. Each sequence within an individual CRISPR is derived from a DNA fragment of a bacteriophage that had previously infected the prokaryote or one of its ancestors (Figure 5). The CRISPR-Cas is RNA interference-based mechanism which protects bacteria from repeated bacteriophage infections by degrading foreign nucleic acids that enter the cell [23].
Our hypothesis: We tried to use some principles of bacterial immunity to suggest possible antigen recognition mechanisms which can function at outright molecular level and doesn’t require pre-existence of billions of immune cells to be prepared for neutralization of any from billions of various antigens existing in Nature.
Prokaryotes can acquire and store information about invading viruses in form of CRISPR sequences. Is it possible that multicellular organisms use similar principles for acquisition and storage of information about invading antigens ? Two known biological phenomena, taken together, can help to answer this question. Namely:
1) Existence of Human Endogenous Retroviruses (HERV) and other transposable elements in our genome [24]; and 2) phenomenon of Defective-Interfering Virus Particles (DIVP) [25]. Thus, we are going to propose a virus-dependent mechanism of acquired immunity.
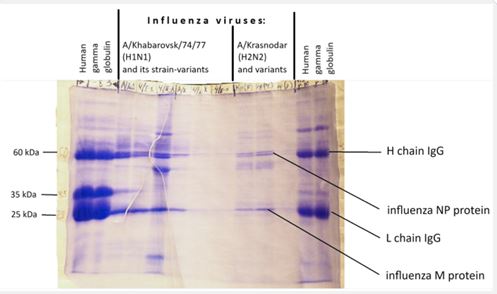
Background: The idea that some unknown viruses or their components may have relation to immunity originated when we performed SDS-PAAG-electrophoresis of influenza viruses and added some samples of human immunoglobulin (Figure 6). The obtained picture showed strange coincidence of electrophoretic mobility between virus proteins and Heavy- and Light IgG chains. From positions of Evolution Theory some interrelation between viruses and immune functions looks quite feasible: The role of viruses as life-initiated entities and drivers of the evolution process is generally appreciated. Therefore it is possible that some virus-adopted and virus-related processes are used in eucariotic organisms, including humans. Perhaps, there’s a reason why some 45% of our DNA is composed of virus-like transposable elements such as LINE, SINE and LTR retrotransposons, including HERVs [26].
The presence of some common features between immunological reactions and virus infection also encourages us to look for possible participation of viruses and virus-like particles in immune reactions (Table 1).
| Similar features | Acquired immunity | Virus infection |
|---|---|---|
| Standard reaction time and staging of the process | Standard timing and staging of antibody production | Standard incubation period, staging and standard timing of clinical symptoms |
| Quick increase of quantity of the involved cells | Clonal expansion of activated immune cells | Increase of quantity of infected cells in geometric progression |
| Shedding from cell membrane (secretion by cell) of specific protein complexes | Secretion of antibodies | Shedding of virus-specific proteins and empty virus capsids from infected cells during aborted infection |
| Expression of specific proteins on the surface of involved cells | Expression of specific antigen receptors on surface of activated immune cells | Clusters of virus-specific proteins on surface of virus- infected cells |
| Identical electrophoretic mobility of definite virus proteins with Ig subunits and other immunity- related proteins | H-chain and L-chain of Ig | NP protein and M protein of influenza virus correspondingly |
| Cytokine production and secretion | Wide spectrum of cytokines | Wide spectrum of cytokines |
| Identical inhibitors | Alkylation, azidothymidine, etc. | Alkylation, azidothymidine, etc. |
Preconditions: Human Endogenous Retroviruses (HERVs)
HERVs are sequences of retroviral origin which represent about 8% of human DNA that is 4 times more than those parts of our genome which encode all human proteins (1 2% of total DNA)! [27]. HERVs sequences have been acquired during millions of years of evolution as the result of multiple integrations by exogenous Retroviruses in the host genome. The retroviral genes encode numerous virus components, including nucleocapsid (gag), Envelope subunits (env), enzymes involved in the viral life cycle: Protease (pro), reverse transcriptase and integrase (pol). HERVs are often coding-defective and thus unable to produce infectious particles [28].
Numerous findings demonstrate possible role of the HERVs in immune reactions: HERVs together with other Retrotransposons (RTP) which constitute about 45% of our genome, can regulate expression of numerous host genes and thus, influence multiple cellular processes, fulfil a role of parallel regulatory network of most vital functions [28-30]. For example, more than one third of p53 binding sites in the human genome are impregnated with HERV sequences, which make HERVs a major component of the p53 regulatory network [31]. Acquisition of new HERVs sequences promote development of important immune pathways and increasing specificity of antiviral defenses [32]. It has been shown that HERV insertions greatly contribute to the function and strengthening of the IFN transcriptional network [29]. This is one more evidence of the significant role of HERVs in innate and adaptive immune responses. Intriguingly, HERVs can produce Env superantigens responsible for polyclonal T-cell activation [33]. Other HERV proteins can significantly regulate gene expression in cells by interaction with thousands of cellular mRNAs [34]. Also of interest is that genetic loci characterized by relatively recent expansion, like those involved in immune reactions, contains more retrotransposons in comparison with more ancient, conserved genes [35]. HERVs are also involved in the development of autoimmune reactions [36].
The most important features of the HERVs supporting their participation in immune reactions: HERVs play significant role in the regulation of host gene expression at several levels: (a) HERVs can trigger chromosomal rearrangements by non-allelic homologous recombination. HERV sequences integrated in proximity to a cellular gene can provide alternative promoters or enhance its expression through LTR regulatory elements. (b) HERV non-coding RNAs (ncRNAs) can regulate cellular genes through recruitment of cellular transcription and splicing factors. HERV ncRNA have been reported to act as “microRNA sponges,” binding and dampening microRNA families responsible for post transcriptional modifications. (c) some HERV proteins can also regulate gene expression through their interaction with cellular mRNAs and the modulation of their transfer and ribosome occupancy [28].
Some HERVs are transmissible inside the host organism, including infection of immune cells [37].
HERVs can induce phenotypic and functional maturation of DC and confer them the potential to support the development of Th1-like effector lymphocytes [38].
HERVs can determine host resistance to viral infection. For example, resistance to infection by MLV in mice is controlled by a Gag-like protein encoded by defective endogenous retrovirus related to human HERV-L [39].
Paleo-genomic investigations demonstrated that origin of adaptive immunity in multicellular organisms coincided in time with unprecedented colonization of their genomes with virus -related sequences which evolved in HERV and other genomic transposable elements [7,32].
One of the most important components of the adaptive immunity, responsible for the self- not self recognition during immune response - the MHC (HLA) complex located on the 6-th chromosome. It is highly polymorphic because it contains insertions of multiple genetic variants of HERVs: concentration of HERVs in this locus of DNA is 10 times higher than in other genome locations that witness direct participation of the HERVs in mechanisms of immune recognition and immune response [40,41].
It is very meaningful that HERV’s proteins have highest expression levels in central immune organ of our body – in thymus: the organ where naïve T lymphocytes acquire their ability to recognize and eliminate antigen-bearing cells. For example, endogenous retrovirus group K Env polyprotein has highest expression in thymocytes but neither expressed nor inducible in mature T-cells [42].
HERVs can be induced by various host and environmental factors, including viral infection: it was shown transactivation of HERVs by a wide range of DNA and RNA viruses that can witness participation of HERVs in defense mechanisms [43].
Preconditions: Defective-Interfering Virus Particles (DIVP)
DIVP - viral particles containing a fraction of the viral genome are able to replicate only in the presence of helper virus and interfere with the intracellular replication of non defective homologous virus. Defective interfering virus particles are a special class of viruses that arise by recombination and rearrangement of viral genomes during replication. DIVPs are defective because they have lost some parts of their genome necessary for replication. Thus, they require presence in the cell (simultaneous infection) of a “helper virus” to provide the function(s) that the defective particle lacks. DI particles have been identified in most virus families [44].
Although the genomes of defective particles are incomplete, they do retain the sequences required for their replication and encapsulation [25].
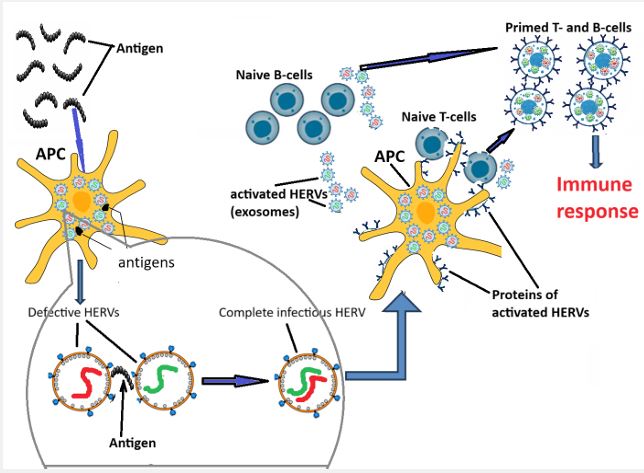
The hypothesis and its discussion: We propose that defective HERVs collectively could ensure recognition of “not self” peptide antigenic determinants trapped by Antigen Presenting Cells (APC). We suggest that attachment of the defective HERV’s components to an antigen inside APC cause reactivation of corresponding clones of HERVs (Figure 7). In the antenatal period reactivated by auto-antigens HERV clones do not quit the APC: instead, they move inside the nucleus and permanently repress DNA of corresponding HERV clones. This happens with participation of the MHC / HLA complexes to prevent auto-immune reactions. As mentioned below, the MHC/HLA - related genomic sequences are hardly impregnated with HERV sequences. These findings provide enormous opportunities for speculation concerning the role of HERVs in the origin, organization and function of the MHC/HLA loci in chromosomes. In the postnatal period all reactivated by binding with antigen clones of the HERVs release from the APC and specifically infect selected types of immune cells (naïve cells), transforming them into clones of B- or T-cells, primed to specific antigen (Table 2).
| Cell clonal selection - based mechanism | Endogenous virus activation - based mechanism (molecular recognition) |
|---|---|
| 1. Extensive heterogenic population of immune cells, bearing Ag receptors of various antigenic specificities preexists in a body; | 1. Extensive heterogenic population of RNA-protein complexes (defective endogenous retroviruses, retrotranspozons, etc.) inhabits (exists) inside APC and other cell populations of a body; |
| 2. Unprimed immune cells have very limited capacity to replicate | 2. The RNA-protein complexes are “defective” i.e., are unable to replicate by themselves; |
| 3. Accidental precise encounter of a cellular receptor with complementary specific antigen triggers mechanism which dramatically increases the replicative capacity of this definite cell clone | 3. Accidental agglomeration of 2 or more RNA-protein complexes (defective endogenous retroviruses, etc.) as a consequence of an entrapped Ag binding, restore the replication ability of this specific type of RNA-protein particles (i.e., transform the specific subpopulation of defective endogenous retroviruses into replicating virus); |
| 4.Negative selection mechanism prevents existence of auto-reactive immune cell clones: preventing of autoimmune reactions; | 4. During embryonic development (i.e., before immune system maturation) thanks to mechanisms of gene silencing and siRNAs inside APC, replication of endogenous defective viruses capable to bind autoantigens is blocked; this prevents presentation of autoantigens by APC; |
| 5. Ag-driven activation of specific immune cell clones (direct or via APC presentation) leads to mounting of the adoptive immune response. | 5. Ag-driven activation of the Ag-binding clones of endogenous retroviruses inside APC initiates robust replication of the specific virus clone and its transmission (in thymus or elsewhere) by exosomes, cell contacts or direct shedding from APC cell membrane to naïve B-and T-cells converting them into primed immune cells capable to mount adoptive immune response. Ag- specific receptors on the immune cell membranes and antibodies represent virus proteins encoded by the activated clone of endogenous retrovirus. |
Proposed mechanism adopts processes and features which are implemented by Nature in other circumstances. For example: 1) to recognize and destroy “not self” RNA in prokaryotes, Nature use CRISPR-Cas nucleic acid -protein complexes; 2) T-lymphotropic retroviruses (like HTLV) are able to selectively infect immune cells causing their activation. 3) Ability of various cells to produce extracellular vesicles - exosomes - has been known for a long time. Exosomes can carry nucleic acids, proteins, lipids, and metabolites, they are mediators of near and long-distance intercellular communication in health and disease and can contribute to various physiological and pathological processes [45,46].
In fact, the mechanism we propose as a machinery of acquired immunity has been already described: it was shown that Gag-like repurposed protein Arc encoded by an ancient retrotransposon is able to mediate communications between neural cells. This happens via assemble of the RTP’s mRNA into capsids to be transferred (distributed) between neurons through extracellular vesicles, then undergoing regulated translation in recipient cells [47]. This plays significant roles in nervous system development. The only difference between this already proven process and our hypothesis of acquired immunity is the necessity of the first preliminary step we postulated, namely: following an antigen binding in an APC by some defective-interfering HERVs, the activation of HERV’s with corresponding “antigen-specific” phenotype should happen; that means initiation of the whole virus or its selected genes transcription and subsequent dissemination between naïve T-and B-cells through virus-like particles = extracellular vesicles. Then in targeted cells synthesis of Ag-binding “repurposed” HERV’s proteins should start, converting them into antigen specific clone of immune cells which displays obtained HERV derived Ag-specific (Ag-complimentary, Ag-binding) proteins on their surface in form of specific Ag receptors and antibodies.
The virus-dependent mechanism of adopted immunity allows to suggest that a mutated reactivated endogenous retrovirus was an ancestor of the HTLV - the etiological agent of AIDS. Because some features of HTLV look like a pathogenic modification of an immunity-related HERV, namely: specific targeting of immune cells (CD4+ lymphocytes) causing their polyclonal activation. In case HTLV would cause monoclonal activation of T-helpers, instead of polyclonal, it could be just a part of the adopted immunity mechanism described above.
Another inference from our proposed Theory could be possibility of the HERV-related mechanism of tumorigenesis: In case reactivated during immune response HERV possesses damaged receptor-binding site, it could attach to unpredicted type of somatic cells, providing them with telomerase activation signal for clonal expansion. This mechanism would explain both kinds of observed correlations: 1) between chronic inflammation and increased threat of neoplasia and 2) between age and incidence of neoplasia. In both cases likelihood of mutations in receptor-binding sites of immunity-related HERVs increasing, thus - increasing probability of their wrong targeting and telomerase activation in a random somatic cell. According to current views, the age-related increase in tumor prevalence is a consequence of accumulation of accidental mutations in somatic cells. But accumulation of accidental mutations would sooner activate apoptosis than telomerase and clonal expansion.
Adoption of the HERV-dependent mechanisms of acquired immunity may promote elaboration of new approaches to prophylaxis and treatment of numerous diseases: autoimmune, oncological and allergic among others. Specifically: reactivation of HERV clones capable of binding autoantigens can be the cause of autoimmune diseases. There are specific gene expression regulators: DNA sequences that function as transcriptional silencers [48]. These silencers can participate in silencing of the “autoreactive” HERV clones to prevent their activation in response to autoantigens. Thus, the cause of autoimmune diseases can be alteration of the autoreactive HERV’s gene silencers because of mutations, or insertion of DNA fragments related to infectious retroviruses or some transposable elements. In this case such treatment approaches could be considered as using HDAC activators, methylating agents or anti-retroviral medicines like RT-inhibitors.
Another implication of the HERV-dependent theory of adaptive immunity can be new vision on predisposing conditions (factors) of allergic disease: for example, shortage of external antigenic impacts can lead to excessive accumulation of naïve T-cells in thymus. Under such conditions, any tiny activation of an immunity-related HERV clone because of antigen binding and expansion of this clone in thymus will result in infection (priming) of excessive number of available naïve immune cells in thymus, that initiate abnormally potent immune response. Under these conditions possible allergy prophylaxis could involve regular training of an individual immune system with new antigens to prevent accumulation of excessive number of naïve T-cells in thymus.
Our principal idea is that all diversity of antigen receptors on the surface of immune cells is not pre-existent but is a result of encounters of every individual immune system with definite restricted number of antigens. It is possible that initial set of antigen receptors in an animal are hereditary obtained from parents who already have met these antigens during their life. This could be possible if oocyte and/or sperm contains some activated, infectious populations of the HERVs.
Therefore, in various individuals (persons) the set of antigen receptors may be slightly or significantly different, depending on the set of antigens they and their parents encountered during their lifespan. The proposed mechanism resembles the CRISPR-Cas defense mechanism in bacteria which also depends on preliminary interaction of an individual bacterial cell (or its ancestor) with a particular virus. With the difference that bacteria incorporate in its genome a piece of virus DNA, while in animal specialized immune cells (in antigen presenting cells - APC and, perhaps, in some other immune cell populations) an antigen activate those pre-existing subpopulations of defective HERVs which bind to an invading antigen. Then activated HERVs infect naive T- and B-cells spreading set of receptor proteins specific for each definite invading antigen.
Thus, we propose the virus-dependent origin of the acquired immunity in animals. We suggest the acquired immunity as a persistent atypical virus infection with following stages: 1) antigen-dependent reactivation of specific clones of the defective HERVs; 2) targeted infection by reactivated HERV clones or their genes of specific naïve immune cells causing their transformation into Ag-specific T- and B-cell clones that is accompanied by shedding of specific HERV proteins from primed B-cells in form of antibodies or their manifestation on cell membrane of primed T-cells. Proposed virus-dependent mechanism of acquired immunity resembles Burnet’s Clonal Selection Theory, but implements antigen recognition on molecular level by virus-like particles (retrotransposons), instead of cell clones: this looks more practical, explains strict timing of immune response and avoids discrepancies during its evaluation from positions of statistics and Probability Theory (Table 2). Importantly that proposed mechanism is much more beneficial from a metabolic point of view because it does not imply waste of energy and resources (proteins, lipids, etc.) on the creation of billions of immune cells specifically targeted against all possible antigens, most of which the animal will never meet during its lifespan.
Abbreviations: Ag: Antigen; APC: Antigen Presenting Cells; Cas: CRISPR-Associated Sequence and CRISPR-Associated Protein (enzyme); CRISPR: Clustered Regularly Interspaced Short Palindromic Repeats; DC: Dendritic Cell; DIVP: Defective Interfering Viral Particles; DNA: Deoxyribonucleic Acid; HERV: Human Endogenous Retrovirus; HLA: Human Leukocyte Antigen Complex; HTLV: Human T-Lymphotropic Virus; IFN: Interferon; LINE: Long Interspersed Nuclear Elements; LTR: Long Terminal Repeats; MHC: Major Histocompatibility Complex; SINE: Short Interspersed Nuclear Elements; TLR: Toll-like Receptor; RTP: Retrotransposons.
Author contributions: AP: Conceptualization, Investigation, Writing-original draft, Literature review, Writing-review & editing VZ: Conceptualization, Review & editing. RJ: Literature review, Validation, editing.
Conflicts of interest: The authors declare that they have no conflicts of interest.
Ethical approval: Not applicable.